1.2.1.
Вегетативная и половая фазы в жизненном
цикле многоклеточных организмов
В клеточных
популяциях прокариотов - бактерий и
синезелёных водорослей - свойство
неограниченной пролиферации присуще всем
клеткам. Это же свойство наблюдается у
простейших, которые относятся к эукариотам.
Прокариоты и простейшие проходят и
жизненном цикле несколько фаз, с которыми
связано изменение их морфологических и
поведенческих свойств. Некоторые
простейшие (например, отдельные
разновидности Sporozoa) в разных фазах
образуют периодически то агрегированную
многоклеточную, то одноклеточную форму [31].
В одну из фаз, связанную с дезагрегацией на
отдельные клетки, образуются не споры, а
половые клетки. У прокариотов и Sporozoa не
образуются субпопуляции
дифференцированных и детерминированных
соматических клеток с ограниченной
пролиферацией, как у более высокоорганизованных
многоклеточных. Дифференцировка у них
происходит циклически: в разных фазах своих
жизненных циклов, а также под влиянием
различных модифицирующих воздействий
внешней среды в клетках развиваются
специальные структуры, характеризующие то
или иное дифференцированное состояние (например,
флагеллы, псевдоподии, оболочки цист,
реснички, раковины и т. д.). Все эти структуры
непостоянны и при смене фаз жизненного
цикла либо при изменениях внешних условий
быстро утрачиваются и сменяются другими.
Важная особенность прокариотов и
простейших та, что все гены их геномов
участвуют в реализации жизненного цикла
[32].
В
многоклеточных организмах ситуация более
сложная. В основе их размножения лежит
особая иммортализованная популяция
клеток, существование которой было открыто
в последнем десятилетии прошлого века |33|. В
1893 г. August Weisman [34] опубликовал «теорию
зародышевой плазмы». Он обратил внимание
на то, что у зародышей животных на разных
стадиях развития обособляется группа
клеток, которые у взрослого организма дают
начало репродуктивным тканям. Эти клетки
были названы автором «непрерывной линией
половых клеток». В настоящее время
существование непрерывной линии половых
клеток подтверждено большим числом
экспериментальных данных [35-37]. Непрерывная
линия половых клеток - это реальный факт
природы, и потому вполне обосновано и
противопоставление друг другу сомы и этой
бессмертной клеточной линии [6].
|
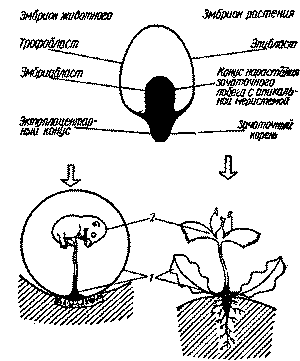
| Рис. 1. Организмы вегетативной и половой фаз плацентарных млекопитающих и высших растений:1 - вегетативная фаза; 2 - половая фаза |
В жизненном
цикле всех многоклеточных эукариотов (включая,
по нашему мнению, и плацентарных
млекопитающих) происходит смена исконных
фаз жизненного цикла - вегетативной и половой.
Вегетативная фаза может завершаться
образованием новых - вегетативных - и
половых организмов (при переходе в половую
фазу). В половой фазе происходит половое
размножение, причем из зиготы всегда
образуется вегетативный организм. Эти
особенности жизненных циклов присущи в
равной мере многоклеточным животным и
растениям (рис. 1). В составе тканей всех
многоклеточных растений есть стойко
воспроизводящая себя субпопуляция
специальных клеток, которая по мере
нормальной пролиферации «выделяет» из
своего состава часть клеток, проходящих
соматическую дифференцировку, образуя
ткани и органы растения. Эту субпопуляцию
клеток в тканях высших растений называют
меристемой [38]. В верхушечной точке роста
побега растения функционирует
Сходным
образом происходит
смена основных фаз
и в жизненном
цикле животных.
Сравнивая различные
виды (многоклеточных)
организмов в соответствующих основных
фазах, мы обнаруживаем ряд общих
морфологических черт: из зиготы образуется
вегетативный организм, причем
морфологически эти возникающие из зиготы
организмы во многом сходны у растений и
животных. Механизм имплантации бластоцисты
животных (образование эктоплацентарного
конуса) напоминает механизм укоренения
семени растений (образование зачаточного
корня) (см. рис. 1 и 2). Остаётся открытым
вопрос, является ли это сходство организмов

 |
|
Рис. 2,
Бластоциста крысы: a - начало имплантации б - стадия двухслойного яйцевого цилиндра; 1 - нмплантационная крипта; 2 - зктоплацентарный конус; 3 - зародышевая эктодерма, 4 - проксимальная ентодерма; 5 - дистальная ентодерма; 6 - трофобласт; 7 - «желточная» полость; 8 -проксимальная ентодерма; 9 -внезародышевая (дистальная) эктодерма; 10 - клетки дистальной эктодермы; 11 - проамниотическая полость 12 - трофоэктодерма |
вегетативных
фаз аналогией или гомологией. В
вегетативной фазе организм, развиваясь,
может пройти от одного до нескольких (а при
соответствующих условиях неограниченное
количество) циклов вегетативного
размножения, и затем образует организм в половой
фазе. При этом образуются гонады, в которые
выселяются предшественники половых клеток.
 |
|
Рис. 3.
Непрерывная линия половых
клеток в жизненном цикле
плацентарных млекопитающих: 1 -выселение гоноцитов из желточного мешка; 2 - мейоз; З - слияние половых клеток с образованием зиготы; 4 - начало дробления зиготы и обособления сомы вегетативной фазы; 5 - период развития организма вегетативной фазы; 6-период закладки сомы организма половой фазы; 7 - период функционирования «апикальной меристемы»; 8 - цикл вегетативного размножения (образование однояйцевых близнецов); 9 - цикл партеногенетического размножения; 10 - момент родов; 11 - период лактации; 12 - завершение полового созревания организма половой фазы; 13 - смерть организма половой фазы; 14 - сома вегетативной фазы; 15 - сома половой фазы; 15 - линия половых клеток |
В составе тканей
организма половой фазы апикальной меристемы
нет, воспроизводство сомы обеспечивается
камбиальными клетками, выделяющими из
своего состава стволовые клетки [39, 40].
Герминативные клетки, заселяющие гонады,
фактически находятся внутри половых
протоков и отграничении от тканей организма
эпителиальным барьером.
Таким образом,
непрерывная линия половых клеток в своем
развитии проходит собственный замкнутый
жизненный цикл (рис. 3), подобный жизненным
циклам прокариотов и Protozoa. В разных фазах
этого цикла происходят временные
дифференцировочные изменения половых
клеток с образованием соответствующих
временных функциональных структур -
псевдоподий, хвостов спермий, zona Pellucida и т. д.
К их числу, по нашему мнению, следует
отнести и сому, для образования которой
жертвуется часть клеток непрерывной линии.
Очевидно, для прохождения непрерывного
цикла в линии половых клеток происходит
периодическая экспрессия - репрессия
определенных групп генов, которые функционируют
только в герминативных клетках. Для клеток,
которые вступают на путь соматической
детерминации, необходима экспрессия еще и
многих других групп генов, которые в линии
половых клеток, по-видимому, никогда не
экспрессируются. Можно предположить, что
последовательность включения генов в
соматических клетках имеет не циклический
характер, как в непрерывной линии половых
клеток, а линейный: от начальной, или инициальной,
группы генов к конечной, или терминальной,
обеспечивающей функцию «рабочей»
смертной соматической клетки. Эта условная
детерминационная линия генной экспрессии
разветвлена в соответствии со "схемой"
дивергенции, благодаря чему образуются
дифференцированные функциональные клетки
разных типов [40].
Главные фазы
жизненного цикла - вегетативная и половая -
у разных животных и растений имеют разный
удельный вес, что проявляется в степени
функциональной дифференцировки сомы
вегетативного либо полового организма,
относительной длительности его
функционирования [18]. Существуют, например,
такие виды метагенетических
кишечнополостных - одиночные или колониальные
полипы, жизнедеятельность которых
протекает преимущественно в вегетативной
фазе [42]. Половые особи (медузоиды) у них
образуются редко, у некоторых колониальных
видов вообще не отделяются от
вегетативной колонии (подобно цветам у
высших растений [12]. Другая крайность -
снижение роли вегетативной фазы и
превалирование половой. Так, сцифоидная медуза
Бугенвиллия (Bougainvillia platygaster) не выбрасывает
яйца в воду. Яйца развиваются у нее прямо в
гонадах, здесь же образуются маленькие
полипы, которые выпочковывают новых маленьких
медуз, а последние отшнуровываютсм и
покидают половые пути материнскою
организма. Среди растений преобладают виды,
у которых главным в жизненном цикле
является вегетативный компонент, но есть и
исключения, например, тропическое
цветковое растение-паразит Раффлезия
Арнольда, у которого побег (вегетативная
фаза) имеется только в раннем зародыше. Из
маленького, с маковое зерно, семени этого
растения появляется сразу маленький
цветочный бутон (половая фаза) без стебля,
который развивается девять месяцев,
достигает размеров большого кочана капусты
и раскрывается в огромный цветок,
достигающий 1 и в диаметре и массой до 6 кг
[43].
Существенное
снижение роли вегетативной фазы произошло
и в жизненном цикле позвоночных животных.
Если некоторые формы примитивных хордовых (туникаты
- сальпы и асцидии) размножаются в
вегетативной фазе не менее эффективно, чем
даже многие кишечнополостные [27], то у
плацентарных млекопитающих вегетативная
фаза проявляет себя в редком явлении рождения
однояйцевых близнецов. Некоторые виды
броненосцев (Dasipodini) всегда рождают парное (от
2 до 12) количество однояйцевых близнецов [37,
44].
Вегетативный
организм в ходе эволюции сначала оказался
помещенным в гонады, затем был заключен в
яйцевую оболочку. У плацентарных
млекопитающих вегетативный организм -
бластоциста - вновь оказался в половых
путях материнского организма, образовав в
зоне контакта плаценту. Вегетативное
размножение в этой фазе стало сравнительно
редким явлением, но оно происходит. Причем,
судя по характеру сращивания близнецовых
плацент, а также появлению иногда
различных сиамских близнецов [45], это
размножение происходит отнюдь не только на
стадии двух бластомеров, как принято
считать, но и на более поздних стадиях
развития организма вегетативной фазы, как
например у броненосцев [37].
По нашему
мнению, факт существования вегетативной и
половой фаз в жизненном цикле животных
является основой для формирования
представлений о биологической сущности
феномена злокачественного роста.